Primats
| Primates | |
|---|---|
 | |
| Període | |
Paleocè – recent | |
| Taxonomia | |
| Super-regne | Eukaryota |
| Regne | Animalia |
| Subregne | Bilateria |
| Fílum | Chordata |
| Classe | Mammalia |
| Superordre | Euarchontoglires |
| Ordre | Primates Linnaeus, 1758 |
| Subordres | |
| |
| Distribució | |
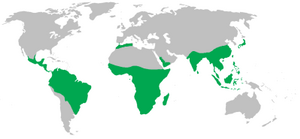 Mapa de distribució dels primats no humans (verd) | |
Els primats (Primates) són un ordre de mamífers que comprèn els lèmurs, els ai-ais, els lorísids, els gàlags, els tarsers, els micos i els simis, amb els humans inclosos en aquesta última categoria.[1] Tret dels humans, que avui en dia habiten tots els continents de la Terra,[a] la majoria de primats viuen a les regions tropicals o subtropicals de les Amèriques, Àfrica i Àsia.[2] La mida dels primats va des dels 30 grams del lèmur ratolí pigmeu fins als 200 quilograms del goril·la de muntanya. Segons les proves fòssils, els avantpassats primitius dels primats podrien haver existit a finals del període Cretaci fa uns 65 milions d'anys i el primatoide més antic conegut és Plesiadapis, del Paleocè superior (fa aproximadament 58-55 milions d'anys). Estudis del rellotge molecular suggereixen que la branca dels primats podria ser encara més antiga i datar del Cretaci mitjà, fa uns 85 milions d'anys.
Tradicionalment s'ha subdividit l'ordre dels primats en dos grups principals: els prosimis i els simis. Els prosimis tenen característiques semblants a les dels primers primats i inclouen els lèmurs de Madagascar, els lorisiformes, els ai-ais i els tarsers. Els simis inclouen els micos i els simis. Més recentment, els taxonomistes han creat el subordre dels estrepsirinis, o primats «de nas moll», per incloure-hi els prosimis no tarsers i el subordre dels haplorins, o primats «de nas sec», per incloure-hi els tarsers i els simiformes. Els simis es divideixen en dos grups: els platirins («de nas pla») o micos del Nou Món, de Sud-amèrica i Mesoamèrica, i els catarins («de nas estret»), d'Àfrica i el sud-est asiàtic. Els micos del Nou Món inclouen els caputxins, els aluates i les mones esquirol i els catarins inclouen els micos del Vell Món (com ara babuïns i macacos) i els simis. Els humans són els únics catarins que s'han estès fora d'Àfrica i el sud-est asiàtic, tot i que les proves fòssils demostren que en el passat també n'existiren moltes espècies a Europa.
Els primats són mamífers generalistes que presenten una gran varietat de característiques. Alguns primats (incloent-hi els humans i els babuïns) no viuen principalment als arbres, però totes les espècies tenen adaptacions per grimpar-hi. Les tècniques de locomoció inclouen saltar d'un arbre a l'altre, caminar sobre dues o quatre potes, caminar sobre els artells, o gronxar-se d'una branca a l'altra (acció coneguda com a braquiació). Els primats es caracteritzen pel seu cervell gran en comparació amb altres mamífers, així com un major ús de la visió estereoscòpica en detriment de l'olfacte, el sistema sensorial dominant en la majoria de mamífers. Aquests trets són especialment importants en els micos i els simis i menys en els loris i els lèmurs. La visió en tres colors ha evolucionat en alguns primats. La majoria tenen polzes oposables i alguns tenen una cua prènsil. Moltes espècies presenten dimorfisme sexual, cosa que significa que els mascles i les femelles tenen trets físics diferents, incloent-hi la massa corporal, la mida dels ullals i la coloració. Els primats tenen un ritme de desenvolupament més lent que altres mamífers de mida similar i assoleixen la maduresa més tard, però tenen una major longevitat. Algunes espècies viuen en solitari, altres en parelles de mascle-femella i altres en grups de fins a centenars de membres.
Contingut
1 Història evolutiva
1.1 Evolució
1.2 Taxonomia
1.3 Híbrids
2 Característiques distintives
3 Anatomia, fisiologia i morfologia
3.1 Dimorfisme sexual
3.2 Locomoció
4 Comportament
4.1 Sistemes socials
4.2 Associacions interespecífiques
4.3 Cognició i comunicació
4.4 Cicle vital
4.5 Dieta i alimentació
5 Hàbitat i distribució
6 Interacció amb els humans
6.1 Estatus legal i social
6.2 Paper en la investigació científica
6.3 Conservació
7 Notes
8 Referències
9 Enllaços externs
Història evolutiva
L'ordre dels primats forma part del clade dels euteris, que es classifica dins el clade dels euarcontoglirs i la classe dels mamífers. Investigacions genètiques moleculars recents sobre els primats, els lèmurs voladors i les tupaies han demostrat que les dues espècies de lèmur volador tenen una relació més propera amb els primats que amb les tupaies,[3] tot i que en el passat les tupaies foren considerades primats.[4] Aquests tres ordres formen el clade dels euarconts. Aquest clade es combina amb el clade dels glirs (compost pels rosegadors i els lagomorfs) per formar el clade dels euarcontoglirs. Segons la font, tant Euarchonta com Euarchontoglires són considerats superordres. D'altra banda, alguns científics consideren que els lèmurs voladors són un subordre dels primats i classifiquen els primats «autèntics» dins el subordre dels euprimats.[5]
Evolució

Plesiadapis, el primatoide més antic
Es creu que el llinatge dels primats es remunta a, com a mínim, fa 65 milions d'anys, tot i que el primatoide més antic conegut del registre fòssil és Plesiadapis (c. 55–58 Ma), que data del Paleocè superior.[6][7] Altres estudis, incloent-hi estudis del rellotge molecular, han estimat que l'origen de la branca dels primats es remunta al període Cretaci mitjà, fa aproximadament 85 milions d'anys.[8][9][10]
En la cladística moderna, l'ordre dels primats és monofilètic. Generalment es creu que el subordre dels estrepsirinis, o «primats de nas moll», divergí de la línia basal dels primats fa uns 63 milions d'anys,[11] tot i que també hi ha indicis que apunten a dates anteriors.[12] Les set famílies d'estrepsirinis són les quatre famílies emparentades de lemuriformes i les tres famílies restants, que inclouen els ai-ais, els lorísids i els gàlags.[13] Alguns sistemes de classificació antics afegeixen els lèmurs mostela als lèmurs i els gàlags amb els lorísids, creant una separació de 3-2 famíles en lloc de la separació 4-3 presentada aquí.[13] Durant l'Eocè, gran part dels continents septentrionals estava dominada per dos grups, els adàpids com i els omòmids.[14][15] Els primers són considerats membres dels estrepsirinis, però mancaven de la pinta dental que tenen els lèmurs moderns. Els últims estaven estretament relacionats amb els tarsers, els micos i els simis. La relació exacta d'aquests dos grups amb els primats vivents és incerta. Els omòmids s'extingiren fa uns 30 milions d'anys,[15] mentre que els adàpids sobrevisqueren fins fa uns 10 milions d'anys.[16]

El lèmur de cua anellada, un primat estrepsirini
Segons estudis genètics, els lèmurs de Madagascar divergiren dels lorisiformes fa aproximadament 75 milions d'anys.[12] Aquests estudis, igual que les proves cromosòmiques i moleculars, també demostren que els lèmurs estan més relacionats ente ells que amb la resta de primats estrepsirinis.[12][17] Tanmateix, Madagascar se separà d'Àfrica fa 160 milions d'anys i de l'Índia fa 90 milions d'anys.[18] Perquè els lèmurs estiguin més relacionats entre ells que amb la resta de primats estrepsirinis, es creu que una minúscula població ancestral arribà a Madagascar en troncs flotants, fa entre 80 i 50 milions d'anys.[12][17][18] S'han examinat altres possibilitats de colonització, com ara múltiples colonitzacions d'Àfrica i Índia, però cap d'elles no compta amb el suport de les proves genètiques i moleculars.[14]
Fins fa poc, l'ai-ai ha sigut difícil de classificar dins els estrepsirinis.[13] S'han proposat teories que la seva família, Daubentoniidae, era o bé un primat lemuriforme (és a dir, que els seus avantpassats se separaren del llinatge dels lèmurs més recentment que la separació entre lèmurs i lorísids) o bé un grup germà de tots els altres estrepsirinis. El 2008, es confirmà que l'ai-ai és un lemuriforme i que descendeix de la mateixa població ancestral de lèmurs que arribà flotant a l'illa. Se'l classifica en l'infraordre dels quiromiformes, formant un clade germà dels lèmurs.[12]
El subordre dels haplorins, o «primats de nas sec», es compon de dos clades germans.[13] Els tarsers prosimis de la família dels tàrsids (monotípica dins del seu propi infraordre dels tarsiformes) representen la divisió més primitiva, fa uns 58 milions d'anys.[19][20] L'infraordre dels simiformes aparegué fa uns 40 milions d'anys,[15] i conté dos clades: el parvordre dels platirins, que es desenvoluparen a Sud-amèrica i inclouen els micos del Nou Món i el parvordre dels catarins, que es desenvoluparen a Àfrica i inclouen els micos del Vell Món, els humans i altres simis.[13] Un tercer clade, que incloïa els eosímids, es desenvolupà a Àsia però s'extingí fa milions d'anys.[21]
Igual que els lèmurs, els micos del Nou Món tenen orígens incerts. Estudis de seqüències moleculars basats en seqüències concatenades han ofert amples variacions en la data estimada de la divergència entre platirins i catarins, que van de fa 70 a fa 33 milions d'anys, mentre que estudis basats en seqüències mitocondrials suggereixen una data més consistent, fa 35 milions d'anys.[22] S'ha suggerit que hi hagué un únic origen dels antropoides a Àfrica i després alguns migraren i es produí una especiació.[14] És possible que els primats antropoides travessessin l'oceà Atlàntic durant l'Eocè, ajudats per dorsals a l'oceà Atlàntic i un nivell del mar més baix i que llavors saltessin d'illa en illa fins a Sud-amèrica.[14] De nou, aquesta colonització transoceànica podria ser explicada perquè els animals hi arribaren en troncs flotants. A causa de la deriva continental, l'oceà Atlàntic jove no era ni de bon tros tan ample com avui en dia,[14] i les investigacions suggereixen que un petit primat d'1 kg podria haver sobreviscut 13 dies en un rai de vegetació.[23] Tenint en compte les velocitats estimades dels corrents i dels vents, podria haver estat un temps suficient per fer el viatge entre els dos continents.

El tití emperador, un mico del Nou Món
Els simis i els micos s'estengueren d'Àfrica a Europa i Àsia a partir del Miocè.[24] Poc després, els loris i els tarsers feren el mateix viatge. Els primers fòssils d'homínids foren descobert al nord d'Àfrica i daten de fa entre 8 i 5 milions d'anys.[15] Els micos del Vell Món desapareguern d'Europa fa uns 1,8 milions d'anys.[25] Els estudis moleculars i dels fòssils mostren generalment que els humans moderns s'originaren a Àfrica fa entre 200.000 i 100.000 anys.[26]
Tot i que els primats estan ben estudiats en comparació amb altres grups d'animals, recentment se n'han descobert algunes espècies noves; proves genètiques efectuades sobre algunes poblacions han revelat algunes espècies anteriorment desconegudes. Primate Taxonomy llistava unes 350 espècies de primats l'any 2001;[27] l'autor, Colin Groves, augmentà aquest nombre fins a 376 en la seva contribució a la tercera edició de Mammal Species of the World (MSW3).[13] Tanmateix, MSW3 es queda curt en relació amb el saber actual, car la col·lecció fou completada el 2003; des d'aleshores, una sèrie de publicacions han pujat aquest nombre fins a 437 espècies. Espècies noves notables no llistades a MSW3 inclouen l'indri de Bemaraha (Avahi cleesei, anomenat en honor de l'actor i entusiasta dels lèmurs britànic John Cleese) i el tití GoldenPalace.com, el nom del qual fou venut en subhasta.
Taxonomia
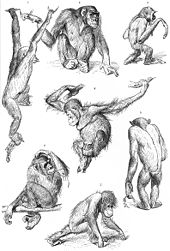
Il·lustració del 1927 de ximpanzés, un gibó (a dalt a la dreta) i dos orangutans (centre i avall al centre). El ximpanzé de dalt a l'esquerra està braquiant, mentre que l'orangutan d'avall al centre camina sobre els artells

Homo sapiens, un membre de l'ordre dels primats

El tarser de les Filipines, antigament considerat un prosimi, ara considerat predominantment un haplorí.
Ordre Primates
Subordre Haplorrhini: tarsers, micos i simis
- Infraordre Simiiformes
- Parvordre Catarrhini
- Superfamília Cercopithecoidea
- Família Cercopithecidae: micos del Vell Món (135 espècies)
- Superfamília Hominoidea
- Família Hominidae: humans i altres grans simis (7 espècies)
- Família Hylobatidae: gibons o «simis inferiors» (13 espècies)
- Superfamília Cercopithecoidea
- Parvordre Platyrrhini: micos del Nou Món
- Família Aotidae: mones de nit (7 espècies)
- Família Atelidae: aluates, mones aranya i mones llanoses (24 espècies)
- Família Cebidae: titís, caputxins i mones esquirol (56 espècies)
- Família Pitheciidae: titís, saquis i uacaris (43 espècies)
- Parvordre Catarrhini
- Infraordre Tarsiiformes
- Família Tarsiidae: tarsers (9 espècies)
- Infraordre Simiiformes
Subordre Strepsirrhini: prosimis no tarsers
- Infraordre Chiromyiformes
- Família Daubentoniidae: ai-ai (1 espècie)
- Infraordre Lemuriformes
- Superfamília Cheirogaleoidea
- Família Cheirogaleidae: lèmurs nans i lèmurs ratolí (33 espècies)
- Superfamília Lemuroidea
- Família Indriidae: indris llanosos i afins (19 espècies)
- Família Lemuridae: lèmurs (22 espècies)
- Família Lepilemuridae: lèmurs mostela (25 espècies)
- Superfamília Cheirogaleoidea
- Infraordre Lorisiformes
- Família Galagidae: gàlags (19 espècies)
- Família Lorisidae: loris, potos i afins (11 espècies)
- Infraordre Chiromyiformes
L'ordre Primates fou creat per Carl von Linné el 1758, a la desena edició del seu llibre Systema Naturae,[28] pels gèneres Homo (humans), Simia (altres simis i micos), Lemur (prosimis) i Vespertilio (ratpenats). A la primera edició del mateix llibre (1735), havia utilitzat el nom Anthropomorpha per Homo, Simia i Bradypus (peresosos).[29] El 1839, Henri Marie Ducrotay de Blainville, seguint Linné i imitant-ne la nomenclatura, creà els ordres Secundates (incloent-hi els subordres Chiroptera, Insectivora i Carnivora), Tertiates (o Glires) i Quaternates (incloent-hi Gravigrada, Pachydermata i Ruminantia),[30] però aquests tàxons nous no foren acceptats.
Abans que Anderson i Jones introduïssin la classificació en estrepsirinis i haplorins el 1984,[31] (seguits per l'obra del 1997 de McKenna i Bell Classification of Mammals: Above the species level),[32] els primats se subdividien en dues superfamílies: els prosimis i els antropoïdeus.[33] Els prosimis incloïen tots els estrepsirinis més els tarsers, mentre que els antropoïdeus incloïen tots els simis.
Híbrids
Els híbrids de primats solen néixer en captivitat,[34] però se n'han donat casos en estat salvatge.[35][36] La hibridació es produeix quan les distribucions de dues espècies s'encavalquen, formant una zona híbrida; els híbrids poden ser creats pels humans quan es posen animals als zoos o a causa de pressions ambientals com ara depredació.[35] També s'han produït hibridacions intergenèriques (híbrids entre membres de gèneres diferents) en estat salvatge. Tot i que pertanyen a gèneres que se separaren fa milions d'anys, encara es produeixen aparellaments entre exemplars de papió gelada i papió sagrat.[37]
Característiques distintives
Els primats s'han diversificat en hàbitats arboris (arbres i matolls) i conserven moltes característiques que són adaptacions a aquest medi.[38] Es distingeixen per:
- la conservació de la clavícula a la cintura escapular;[38]
articulacions glenohumerals que permeten un alt grau de moviment en totes les direccions;[38]
- cinc dits a cada pota, amb polzes oposables i dits dels peus grossos;[38]
- ungles als dits de les mans i els peus hola soc molt guapa;[39]
- una ungla plana a l'hàl·lux (en totes les espècies vivents);[39]
- coixinets tàctils sensibles a la punta dels dits;[38]
òrbites envoltades per os;[40]
- una tendència vers un musell reduït i una cara aplanada, atribuïda a una confiança en la vista en detriment de l'olfacte (especialment en els haplorins i menys en els estrepsirinis);[40]
- un sistema visual complex amb visió esteoroscòpica, una elevada agudesa visual i visió en color;[38]
- un cervell amb un cerebel ben desenvolupat dotat d'un lòbul posterior i una cissura calcarina;[40]
- un cervell gran en proporció a la mida corporal, especialment en els simiformes;[38]
- una diferenciació d'una escorça cerebral agrandida;[38]
- un nombre reduït de dents en comparació amb els mamífers primitius;[38]
- tres tipus de dents;[40]
- un cec ben desenvolupat;[40]
- dues glàndules mamàries pectorals;[38]
- habitualment, una cria per gestació;[38]
- un penis pèndol i testicles escrotals;[40]
- un període de gestació i de desenvolupament llarg;[38] i
- una tendència vers mantenir el tors erecte que condueix al bipedisme.[38]
No tots els primats presenten aquests trets anatòmics, ni tots els trets són únics a aquests animals. Per exemple, altres mamífers tenen clavícules, tres tipus de dents i un penis pèndol, mentre que les mones aranya tenen polzes molt reduïts, els lèmurs de collar tenen sis glàndules mamàries i els estrepsirinis solen tenir un musell més llarg i un gran sentit de l'olfacte. Els primats són mamífers generalistes.[40]
Quant al comportament, els primats són animals altament socials, amb jerarquies de dominància flexibles.[41] Les espècies del Nou Món formen lligams de parella monògams i presenten una cura paterna significativa de les seves cries, a diferència de la majoria de micos del Vell Món.[42]
Anatomia, fisiologia i morfologia
Els primats tenen ulls que miren cap endavant a la part anterior del crani; la seva visió binocular els permet estimar la distància amb precisió, cosa útil pels avantpassats braquiants dels humans.[38] Hi ha una cresta òssia a sobre de les òrbites oculars; aquesta cresta reforça ossos més dèbils de la cara que es distenen durant la masticació. Els estrepsirinis tenen una barra postorbitària, un os que envolta l'òrbita ocular, per protegir-se els ulls; en canvi, els primats superiors, els haplorins, han desenvolupat òrbites completament tancades.[43]

Mida i comparació dels cranis de primats.
El cap dels primats té un gran crani voltat que és especialment prominent en els antropoides. El crani protegeix un cervell gran, que és una característica distintiva d'aquest grup.[38] El volum endocranial (el volum dins del crani) és tres vegades superior en els humans que en el primat no humà més gran, reflectint una major mida del cervell.[44] El volum endocranial mitjà és de 1.201 centímetres cúbics en els humans, 469 cm³ en els goril·les, 400 cm³ en els ximpanzés i 397 cm³ en els orangutans.[44] La tendència evolutiva principal dels primats ha sigut el desenvolupament del cervell, en particular el neocòrtex (una part de l'escorça cerebral), que té un paper en la percepció sensorial, la generació d'ordres motrius, el raonement espacial, el pensament conscient i, en els humans, el llenguatge.[2] Mentre que altres mamífers confien molt en el seu olfacte, la vida arborícola dels primats ha conduït a un sistema sensorial dominat pel tacte i la vista,[2] una reducció de la regió olfactiva del cervell i un comportament social cada vegada més complex.[45]

Il·lustració del 1893 de les mans i els peus de primats.
Els primats tenen generalment cinc dits a cada pota (pentadactília), amb ungles de ceratina a l'extrem de cada dit. La part inferior de les mans té coixinets sensibles a les polpes dels dits. La majoria tenen polzes oposables, un tret característic dels primats; tanmateix, els polzes oposables no estan limitats a aquest ordre (els opòssums, per exemple, també en tenen).[38] Els polzes permeten a algunes espècies utilitzar eines. En els primats, la combinació de polzes oposables, ungles curtes (en lloc d'urpes) i llargs dits que es tanquen cap endins és una relíquia de la pràctica ancestral d'agafar-se a branques i, en part, ha permès a algunes espècies desenvolupar la braquiació (gronxar-se d'una branca a l'altra agafats pels braços) com a mitjà de locomoció significatiu. Els prosimis tenen unes ungles semblants a urpes al segon dit de cada peu, anomenades urpes d'empolainament, que fan servir per empolainar-se.[38]
La clavícula dels primats és conservada com a un dels elements prominents de la cintura escapular; això permet una àmplia mobilitat de l'articulació glenohumeral (l'articulació de l'espatlla).[41] Els simis tenen articulacions glenohumerals i braços més mòbils gràcies a la posició dorsal de l'escàpula, una caixa toràcica que és més plana en el pla frontal-dorsal i una columna vertebral més curta i menys mòbil en comparació amb els micos del Vell Món (amb una gran reducció de les vèrtebres inferiors que en algunes espècies ha conduït a la pèrdua de la cua). Els micos del Vell Món es diferencien dels simis pel fet que la majoria tenen cua. L'única família de primats amb una cua prènsil són els atèlids del Nou Món, que inclouen els aluates, les mones aranya i les mones llanoses.
Els primats mostren una tendència evolutiva vers un musell reduït.[41] Tècnicament, es distingeixen els micos del Vell Món dels micos del Nou Món per l'estructura del nas i es distingeixen dels simis per la configuració de les seves dents.[45] En els micos del Nou Món, els narius apunten als costats, mentre que en els del Vell Món, apunten cap avall.[45] Els primats tenen una configuració dental considerablement variada i tot i que alguns han perdut la majoria de les incisives, tots conserven com a mínim una incisiva inferior.[45] En la majoria d'estrepsirinis, les incisives i canines inferiors formen una pinta dental, que s'utilitza per a l'empolainament i per buscar aliment,[40][45] i la primera premolar inferior té forma de canina.[40] Els micos del Vell Món tenen vuit premolars, mentre que els del Nou Món en tenen dotze.[45] Les espècies del Vell Món es divideixen en simis i micos segons el nombre de cúspides que tinguin a les dents molars; els simis en tenen cinc i els micos en tenen quatre,[45] tot i que els humans poden tenir-se quatre o cinc.[46] La cúspide molar principal dels homínids (l'hipocon) evolucionà al principi de la història dels primats, mentre que la cúspide de la molar inferior primitiva corresponent (el paracònid) es perdé. Els prosimis es distingeixen pels seus llavis superiors immobilitzats, la punta humida del seu nas i les incisives inferiors que apunten cap endavant.
L'evolució de la visió en color en els primats és única entre la majoria de mamífers euteris. Mentre que els avantpassats llunyans vertebrats dels primats tenien una visió en tres colors (tricromàcia), els seus avantpassats mamiferoides nocturns i de sang calenta perderen un dels tres cons de la retina durant el període Mesozoic. Per tant, els peixos, els rèptils i els ocells són tricromàtics o tetracromàtics, mentre que tots els mamífers, tret d'alguns primats i marsupials,[47] són dicromàtics o monocromàtics (no veuen els colors).[40] Els primats nocturns, com ara les mones de nit i els gàlags, solen ser monocromàtics. Els catarins són generalment tricromàtics gràcies a una duplicació del gen d'opsines vermell-verd a la base del seu llinatge, fa entre 40 i 30 milions d'anys.[40][48] Els platirins, d'altra banda, són rarament tricromàtics.[49] Concretament, algunes femelles han de ser heterozigòtiques per dos al·lels del gen d'opsines (vermell i verd) situat al mateix locus del cromosoma X.[40] Així doncs, els mascles només poden ser dicromàtics, mentre que les femelles poden ser o bé dicromàtiques o bé tricromàtiques. La visió en color en els estrepsirinis no està ben comprès; tanmateix, les investigacions indiquen una varietat de visió en color semblant a la dels platirins.[40]
Com els catarins, els aluates (un grup de platirins) tenen una tricromàcia generalitzada que es remunta a una duplicació genètica evolutivament recent.[50] Els aluates són uns dels folívors més especialitzats dels micos del Nou Món; les fruites no són una part important de la seva dieta,[51] i el tipus defulles que prefereixen consumir (joves, nutritives i digeribles) només es poden detectar per un senyal vermell-verd. El treball de camp sobre les preferències alimentàries dels aluates suggereix que la tricromàcia generalitzada fou afavorida per la selecció ambiental.[49]
Dimorfisme sexual

Es pot observar un clar dimorfisme sexual de mida entre el papió sagrat mascle (gris) i la femella (marró).
El dimorfisme sexual, la variació entre individus de diferent sexe dins la mateixa espècie, és sovint present en els simis, tot i que en major mesura en les espècies del Vell Món (simis i alguns micos) que les del Nou Món. Estudis recents utilitzen la comparació d'ADN per examinar tant la variació en l'expressió del dimorfisme entre els primats com les causes fonamentals del dimorfisme sexual. Els primats solen tenir dimorfisme de massa corporal[52][53] i de mida dels ullals[54][55] així com del color del pelatge i la pell.[56] El dimorfisme es pot atribuir i és afectat per diferents factors, incloent-hi el sistema d'aparellament,[57] la mida,[57] l'hàbitat i la dieta.[58]
Les anàlisis comparatives han generat una comprensió més completa de la relació entre la selecció sexual, la selecció natural i els sistemes d'aparellament dels primats. Els estudis han demostrat que el dimorfisme és el resultat de canvis en els trets tant dels mascles com de les femelles.[59] La dilatació ontogenètica, en què es produeix una extensió relativa d'una trajectòria de creixement comuna, podria oferir informació sobre la relació entre el dimorfisme sexual i els patrons de creixement.[60] Algunes proves del registre fòssil suggereixen que hi hagué una evolució convergent del dimorfisme i alguns homínids extints probablement tenien un dimorfisme més marcat que qualsevol primat vivent.[59]
Locomoció

Un sifaca de diadema, que s'aferra a les branques i salta d'una a l'altra, es prepara per saltar a un altre arbre.
Les espècies de primats es mouen per braquiació, bipedisme, salts, quadrupedisme arborícola i terrestre, grimpant, caminant sobre els artells, o per una combinació d'aquests mètodes. Alguns prosimis es desplacen principalment grimpant i saltant per les branques. En són exemples els gàlags, els índrids (com els sifaques, els indris llanosos o els indris), els lèmurs mostela i tots els tarsers.[61] Altres prosimis són quadrúpedes i grimpadors arborícoles. Alguns també són quadrúpedes terrestres, mentre que d'altres són saltadors. La majoria de micos són quadrúpedes i grimpadors tant terrestres com arborícoles. Els gibons i les mones aranya llanoses i les mones aranya utilitzen sovint la braquiació.[25] Les mones llanoses també braquien de tant en tant.[51] Els orangutans utilitzen una forma de locomoció similar anomenada escalada quadrumana, en què utilitzen els braços i les cames per moure el seu cos pesant pels arbres.[25] Els ximpanzés i els goril·les caminen sobre els seus artells,[25] i es poden moure a dues potes en distàncies curtes. Tot i que nombroses espècies, com ara els australopitecins i els homínids primerencs, han presentat una locomoció completament bípeda, els humans són l'única espècie vivent amb aquesta característica.
Comportament
Sistemes socials
Richard Wrangham afirmà que la millor manera de classificar els sistemes socials dels primats no humans és segons la freqüència amb què les femelles canvien de grup.[62] Proposà quatre categories:
- Sistemes de transferència de femelles: les femelles abandonen el grup en què han nascut. Les femelles d'un grup no tindran una relació propera, mentre que els mascles romanen amb el seu grup natal i aquesta estreta associació pot influir en el comportament social. Els grups solen ser bastant petits. Es pot observar aquest sistema en els ximpanzés, en què els mascles, que típicament estan relacionats, cooperen en la defensa del territori del grup. Entre els micos del Nou Món, les mones aranya i les mones aranyes llanoses utilitzen aquest sistema.[63]

Uns macacos del Japó es banyen ensems a les Aigües Termals de Jigokudani.
- Sistemes de transferència de mascles: mentre les femelles romanen al seu grup natal, els masclen emigren en arribar a l'adolescència. Les societats poligines i multi-mascles es classifiquen dins aquesta categoria. La mida del grup sol ser més gran. Aquest sistema és habitual entre els lèmurs de cua anellada i els caputxins, així com en els cercopitecins.[25]
- Espècies monògames: un lligam mascle-femella, a vegades acompanyat per les cries joves. Es comparteixen la responsabilitat de la cura parental i la defensa del territori. Les cries abandonen el territori dels pares durant l'adolescència. Els gibons utilitzen essencialment aquest sistema, tot i que en aquest context «monogàmia» no significa necessàriament fidelitat sexual absoluta.[64]
- Espècies solitàries: sovint són mascles que defensen territoris que inclouen el territori de diverses femelles. Aquest tipus d'organització es dóna en els prosimis. Els orangutans no defensen el seu territori però a la pràctica tenen aquest tipus d'organització.[65]
També es coneixen altres sistemes. Per exemple, en els aluates tant els mascles com les femelles solen abandonar el seu grup natal per un altre quan assoleixen la maduresa sexual, amb el resultat que hi ha grups en què en general ni els mascles ni les femelles estan relacionats.[51] Alguns prosimis, micos colobins i micos cal·litríquids utilitzen aquest sistema.[25]

Els ximpanzés són animals socials.
La primatòloga Jane Goodall, que estudià al Parc Nacional de Gombe Stream, observà societats de fissió-fusió en els ximpanzés.[66] Es produeix «fissió» quan el grup principal es dispersa per recollir aliments durant el dia i «fusió» quan el grup es reuneix de nit per dormir tots junts. Aquesta estructura social també es pot observar en el papió sagrat,[67] les mones aranya[51] i els bonobos.[67] El papió gelada té una estructura social similar en què nombrosos grups petits es reuneixen per formar grups temporals de fins a 600 micos.[67]
Aquests sistemes socials estan afectats per tres factors ecològics principals: la distribució dels recursos, la mida del grup i la depredació.[42] Dins d'un grup social hi ha un equilibri entre la cooperació i la competició. Els comportaments cooperatius inclouen l'empolainament social (treure's paràsits de la pell i netejar-se ferides mútuament), compartir aliments i defensar-se col·lectivament dels predadors o d'invasors. Els comportaments agressius solen implicar competició pels aliments disponibles, pels llocs per dormir o per parelles sexuals. L'agressió també es fa servir per establir jerarquies de dominància.[42]
Associacions interespecífiques
Se sap que algunes espècies de primats s'associen en estat salvatge. Algunes d'aquestes associacions han estat estudiades en detall. Al Parc Nacional de Taï (Àfrica), diverses espècies coordinen el seu comportament anti-predadors. En són exemples el cercopitec diana, el cercopitec de Campbell, el cercopitec de nas blanc petit, el còlob vermell occidental, el còlob de crinera occidental i el mangabei de collar, que coordinen les seves crides d'alarma anti-predadors.[68] El ximpanzé comú és un dels predadors d'aquests micos.[69]
El cercopitec de cua vermella s'associa amb diverses espècies, incloent-hi el còlob vermell occidental, el cercopitec blau, el cercopitec de Wolf, el còlob de crinera oriental, el mangabei negre i el cercopitec d'Allen.[67] Diverses d'aquestes espècies són preses del ximpanzé comú.[70]
A Sud-amèrica, les mones esquirol s'associen amb els caputxins.[71] Això pot tenir més a veure amb els beneficis alimentaris que no pas amb beneficis anti-predadors.[71]
Cognició i comunicació
Els primats tenen capacitats cognitives avançades: alguns fabriquen eines i les utilitzen per adquirir aliment o en exhibicions socials;[72][73] d'altres tenen estratègies de caça sofisticades que requereixen cooperació, influència i rangs;[74] són conscients del seu estatus, manipuladors i capaços d'enganyar;[75] poden reconèixer els parents i els coespecífics;[76][77] i poden aprendre a utilitzar símbols i entendre aspectes del llenguatge humà, incloent-hi una certa sintaxi relacional i conceptes numèrics, així com seqüències numèriques.[78][79][80] La investigació de la cognició dels primats explora la resolució de problemes, la memòria i conceptes numèrics, espacials i abstractes.[81]
Els lèmurs, lorísids, tarsers i micos del Nou Món confien en els senyals olfactius per molts aspectes del seu comportament social i reproductiu.[2] Utilitzen glàndules especials per marcar territori amb feromones, que són detectades per l'òrgan vomeronasal; aquest procés forma una part important del comportament comunicatiu d'aquests primats.[2] En els micos del Vell Món i els simis, aquesta capacitat és en gran part vestigial, havent regredit a mesura que els ulls tricromàtics evolucionaven per convertir-se en l'òrgan sensorial principal.[82] Els primats també utilitzen vocalitzacions, gestos i expressions facials per comunicar el seu estat psicològic.[83]
Cicle vital
Els primats tenen taxes de desenvolupament més lentes que altres mamífers.[25] Tots els infants de primats no humans són alletats per la seva mare i en depenen per l'empolainament i el transport.[25] En algunes espècies, els infants són protegits i transportats per mascles del grup, particularment els que poden ser els seus pares.[25] Altres parents de l'infant, com ara germans i tietes, també poden participar a cuidar-lo.[25] La majoria de primats mares deixen d'ovular mentre alleten un infant; una vegada l'infant és deslletat, la mare es pot reproduir de nou.[25] Això sol provocar un conflicte de deslletament amb els infants que intenten continuar mamant.[25]
Els primats tenen un període juvenil més llarg entre el deslletament i la maduresa sexual que altres mamífers de mida similar.[25] Durant el període juvenil, els primats són més susceptibles que els adults a la predació i la inanició; durant aquest temps, adquireixen experiència en alimentar-se i evitar els predadors.[25] Adquireixen habilitats socials i de combat, sovint per mitjà de jocs.[25]
Els primats, especialment les femelles, tenen una major longevitat que altres mamífers de mida similar.[25]
Dieta i alimentació

Un còlob de crinera oriental, una espècie de còlob, s'alimenta de fulles.
Els primats exploten una varietat de fonts d'aliment. La majoria de primats inclouen fruita a la seva dieta per obtenir glúcids i lípids fàcils de digerir com a font d'energia.[25] Tanmateix, necessiten altres aliments, com ara fulles o insectes, per obtenir aminoàcids, vitamines i minerals. Molts primats tenen especialitzacions anatòmiques que els permeten explotar aliments concrets, com ara fruita, fulles, resina o insectes.[25] Per exemple, els folívors com ara els aluates, els còlobs i els lèmurs mostela tenen un tracte digestiu estès que els permeten absorbir nutrients de fulles que poden resultar difícils de digerir.[25] Els titís, que s'alimenten de resina, tenen dents incisives fortes, que els permeten obrir l'escorça dels arbres per arribar a la resina i tenen urpes en lloc d'ungles, cosa que els permet agafar-se als arbres mentre s'alimenten.[25] L'ai-ai combina dents semblants a les dels rosegadors amb un llarg i prim dit del mig per omplir el mateix nínxol ecològic que els picots. Pica als arbres per detectar larves d'insecte, rosega forats a la fusta i hi insereix el seu allargat dit del mig per treure'n les larves.[84] Algunes espècies tenen especialitzacions addicionals. Per exemple, el mangabei de galtes grises té un espès esmalt a les dents, que li permet obrir fruits i llavors durs que els altres micos no poden obrir.[25]
El papió gelada és l'única espècie de primat que s'alimenta principalment d'herba.[85] Els tarsers són els únics primats carnívors, alimentant-se exclusivament d'insectes, crustacis, petits vertebrats i serps (incloent-hi espècies verinoses).[86] Els caputxins, d'altra banda, poden utilitzar molts tipus diferents de menjar, incloent-hi fruits, fulles, flors, borrons, nèctar, llavors, insectes i altres invertebrats, ous d'ocell i petits vertebrats com ara ocells, sargantanes, esquirols i ratpenats.[51] El ximpanzé comú té una dieta variada que inclou la depredació d'altres espècies de primat, com ara el còlob vermell occidental.[69][70]
Hàbitat i distribució

Macaco rhesus al Fort d'Agra (Índia).
Els primats evolucionaren d'animals arborícoles i la majoria d'espècies passen gran part de la vida als arbres. Moltes espècies de primats viuen en selves tropicals. S'ha demostrat que el nombre d'espècies de primats a les àrees tropicals està positivament correlacionat amb la quantitat de precipitacions i l'àrea de superfície de la selva.[87] Els primats representen entre un 25% i un 40% dels animals frugívors (en pes) de les selves tropicals i tenen un paper ecològic important dispersant les llavors de moltes espècies d'arbres.[88]
Alguns animals són parcialment terrestres, com ara els papions i les mones vermelles i algunes espècies són completament terrestres, com ara el papió gelada i els humans. Els primats no humans viuen en una gran varietat d'hàbitats forestats de les latituds tropicals d'Àfrica, l'Índia, el sud-est asiàtic i Sud-amèrica, incloent-hi les selves pluvials, els boscos de manglars i els boscos de montana. Hi ha alguns exemples de primats no humans que viuen a l'exterior dels tròpics; el macaco del Japó, que viu a les muntanyes, habita el nord de Honshū, on hi ha neu vuit mesos a l'any; la mona de Gibraltar viu a la serralada de l'Atles d'Algèria i el Marroc. Els hàbitats dels primats abasten una varietat d'altituds: el rinopitec daurat ha sigut trobat vivint al Hengduan Shan, a altituds de 4.700 metres,[89] el goril·la de muntanya es pot trobar a 4.200 metres d'altitud, travessant les muntanyes Virunga,[90] i s'ha trobat el papió gelada a elevacions de fins a 5.000 metres al massís etíop. Tot i que a la majoria d'espècies no els agrada l'aigua, algunes són bones nedadores i es troben còmodes en aiguamolls i zones aquoses, incloent-hi el nassut, el cercopitec de Brazza i el cercopitec d'Allen, que ha desenvolupat unes petites membranes entre els dits. Alguns primats, com el macaco rhesus i els langurs, poden explotar medis modificats pels humans i fins i tot viure en ciutats.[67][91]
Interacció amb els humans
Alguns han plantejat la hipòtesi que la suposada relació estreta les interaccions (a vegades sexuals[92]) entre els humans i els primats no humans creen rutes per la transmissió d'agents causants de malalties zoonòtiques. Els virus com els herpesvírids (notablement l'herpesvirus B), els poxvírids, el xarampió, l'ebola, la ràbia, el virus de Marburg i l'hepatitis vírica es poden transmetre als humans; en alguns casos els virus produeixen malalties potencialment fatals tant als humans com als primats no humans.[93]
Estatus legal i social
Només els humans són reconeguts com a persones i estan protegits per la llei, per la Declaració Universal dels Drets Humans de les Nacions Unides.[b] L'estatus legal dels primats no humans (PNH), d'altra banda, és objecte d'un intens debat i hi ha organitzacions com ara el Projecte Gran Simi (PGS, o GAP per les seves sigles en anglès) que fan campanya perquè s'atorgui als primats almenys alguns drets legals.[94] Al juny del 2008, Espanya esdevingué el primer país del món en reconèixer els drets d'alguns PNH quan el comitè ambiental interpartidista del seu parlament incità l'estat a complir les recomanacions del PGS, que són que els ximpanzés, els bonobos, els orangutans i els goril·les no siguin utilitzats per l'experimentació animal.[95][96]

Un home pidola diners ensenyant un primat exòtic als turistes.
Moltes espècies són tingudes com a animals de companyia pels humans. El PGS estima que uns 3.000 PNH viuen com a animals de companyia exòtics als Estats Units, mentre que la Humane Society of the United States dóna una xifra molt més elevada, d'uns 15.000.[97] La creixent classe mitjana xinesa ha incrementat la demanda de PNH com a animals de companyia exòtics als últims anys.[98] Tot i que la importació de PNH com a animals de companyia fou prohibida el 1975 als Estats Units, encara es fa contraban a la frontera EUA-Mèxic, amb preus que van de 3.000$ per un mico fins a 30.000$ per un simi.[99]
Els primats són utilitzats com a organismes model en laboratoris i se'ls ha utilitzat en missions espacials.[100] Serveixen d'animals de servei pels humans discapacitats. Es poden ensinistrar micos caputxins perquè ajudin els humans tetraplègics; la seva intel·ligència, memòria i destresa manual en fan ajudants ideals.[101]

La destresa manual dels caputxins és un dels motius pels quals poden ajudar els humans tetraplègics.
Es tenen PNH en zoològics d'arreu del món. Històricament, els zoos han estat principalment una forma d'entreteniment, però més recentment s'han centrat en la conservació, l'educació i la investigació. Avui en dia, molts zoos tenen exhibicions naturalistes i material educatiu pel públic; als Estats Units, molts participen en el Species Survival Plan (SSP), desenvolupat per l'Associació de Zoos i Aquaris (AZA), per maximitzar la diversitat genètica per mitjà de la cria en captivitat. Els zoos i altres paladins del benestar animal solen oposar-se a la iniciativa dels grups pels drets dels animals i a la insistència del PGS que tots els PNH siguin posats en llibertat per dues raons principals. Primerament els primats nascuts en captivitat manquen dels coneixements i l'experiència necessaris per sobreviure en estat salvatge si són alliberats. Segonament, els zoos ofereixen un espai vital pels primats i altres animals que en llibertat estan amenaçats d'extinció.
Paper en la investigació científica
S'utilitzen milers de primats no humans arreu del món en la investigació a causa de la seva suposada semblança psicològica i fisiològica amb els humans.[102][103] En particular, el cervell i els ulls dels PNH són més similars als dels humans que els de qualsevol altre animal. Els PNH són utilitzats habitualment en assajos preclínics, la neurociència, estudis d'oftalmologia i estudis de toxicitat. Els macacos rhesus són utilitzats sovints, igual que altres macacos, les mones verdes, els ximpanzés, els papions, les mones esquirol i els titís, tant capturats del seu hàbitat natural com criats a propòsit.[102][104] El 2005, el PGS informà que 1.280 dels 3.100 PNH que vivien en captivitat als Estats Units eren utilitzats per experiments.[94] El 2004, la Unió Europea utilitzà uns 10.000 PNH en aquest tipus d'experiments; el 2005, a Gran Bretanya es dugueren a terme 4.652 experiments sobre 3.115 PNH.[105] Els governs de molts països tenen unes regles estrictes per la cura dels PNH tinguts en captivitat. Als Estats Units, les regles federals regulen detalladament aspectes de l'allotjament, l'alimentació, l'enriquiment i la cria dels PNH.[106] Grups europeus com ara la Coalició Europea per a l'Abolició dels Experiments amb Animals tenen com a objectiu la prohibició d'utilitzar PNH en experiments com a part de la revisió de la Unió Europea de la seva legislació sobre la investigació animal.[107]
Conservació
La Unió Internacional per a la Conservació de la Natura (UICN) compila cada dos anys la llista dels 25 primats en major perill d'extinció, que va a càrrec del Primate Specialist Group i la Species Survival Commission de la UICN i la Societat Primatològica Internacional, en col·laboració amb Conservation International i la Bristol Conservation and Science Foundation.
L'edició del 2012 contenia nou espècies d'Àsia, sis de Madagascar, cinc de l'Àfrica continental i cinc de l'àmbit neotropical. En termes de països, Madagascar encapçala la llista amb sis espècies, mentre que el Vietnam en té cinc, Indonèsia tres, el Brasil dues i la Xina, Colòmbia, la Costa d'Ivori, la República Democràtica del Congo, l'Equador, la Guinea Equatorial, Ghana, Kenya, el Perú, Sri Lanka, Tanzània i Veneçuela una espècie cadascun.
Hi destaquen espècies com ara el tarser pigmeu, del qual només es coneixien tres espècimens conservats a museus fins que el 2008 en foren capturats tres individus al Parc Nacional Lore Lindu i se n'observà un altre de silvestre. Els lèmurs de Madagascar estan molt afectats per la destrucció d'hàbitat i la caça il·legal. El lèmur mostela septentrional és el més rar de tots, amb només dinou individus en llibertat.
Les amenaces més comunes inclouen la desforestació, la fragmentació forestal i l'extermini de micos (el resultat de les ràtzies dels conreus que duen a terme els primats),[108] així com la caça de primats per utilitzar-los com a medicaments, animals de companyia o aliment. La destrucció de boscos tropicals a gran escala és àmpliament vista com el procés que més amenaça els primats.[109][110][111] Més del 90% d'espècies de primats viuen als boscos tropicals.[110][112] La causa principal de la pèrdua de boscos és la desforestació per fer lloc a conreus, tot i que la indústria fustera, la recol·lecció de subsistència de fusta, la minera i la construcció de preses també contribueixen a la destrucció dels boscos.[112]

L'orangutan de Sumatra està en perill crític
Els primats de gran mida (més de 5 kg) corren un major risc d'extinció a causa de la seva major rendibilitat pels caçadors furtius, en comparació amb els primats més petits.[112] Assoleixen la maduresa sexual més tard que altres animals i tenen un període més llarg entre parts. Per tant, les poblacions tenen menys temps de recuperació després d'haver perdut membres a la caça furtiva o el comerç d'animals de companyia.[113] Les dades d'algunes ciutats africanes mostren que la meitat de les proteïnes consumides en àrees urbanes prové del comerç en bushmeat.[114] Primats amenaçats com ara cercopitecs i els drils són caçats a una escala que supera de llarg els nivells sostenibles.[114] Això es deu a la seva mida corporal gran, facilitat de transport i rendibilitat per animal.[114] Com que l'agricultura envaeix els hàbitats forestals, els primats s'alimenten dels conreus, provocant grans pèrdues econòmiques als grangers.[115] Les ràtzies dels conreus que duen a terme els primats fa una impressió negativa dels primats, perjudicant els esforços de conservació.[116]
Madagascar, que té cinc famílies de primats endèmiques, ha experimentat l'extinció més gran del passat recent; des que els humans colonitzaren l'illa fa 1.500 anys, almenys vuit classes i quinze espècies s'han extingit a causa de la caça i la destrucció d'hàbitat.[2] Entre els primats que s'extingiren hi havia Archaeoindris (un lèmur més gran que un goril·la mascle adult) i les famílies dels paleopropitècids i arqueolemúrids.[2]
A Àsia, l'hinduisme, el budisme i l'islam prohibeixen el consum de carn de primat; tanmateix, els primats són caçats com a aliment de totes maneres[112] Algunes religions tradicionals permeten el consum de carn de primat.[117][118] El comerç d'animals de companyia i la medicina tradicional també incrementen la demanda per la caça il·legal.[98][119][120] El macaco rhesus, un organisme model, fou protegit després que la captura excessiva n'amenacés les poblacions a la dècada del 1960; el programa fou tan eficaç que els macacos són actualment considerats una plaga arreu del seu àmbit de distribució.[111]

El goril·la del riu Cross és una espècie en perill crític.
A Mesoamèrica i Sud-amèrica, la fragmentació forestal i la caça són els dos problemes principals pels primats. Actualment, les grans zones boscoses són rares a Mesoamèrica.[109][121] Això augmenta la quantitat de bosc vulnerable a efectes de vora com ara la invasió de conreus, nivells més baixos d'humitat i un canvi en la vida vegetal.[122][123] La restricció dels moviments resulta en una major quantitat d'endogàmia, que pot causar efectes deleteris que condueixen a un coll d'ampolla poblacional, en què es perd un percentatge significatiu de la població.[124][125]
Hi ha 21 primats en perill crític, vuit dels quals han estat a la llista dels 25 primats més amenaçats del món de la UICN des de l'any 2000: el sifaca sedós, el langur de François, el langur de cap daurat, el rinopitec de potes grises, el rinopitec de Tonquín, Nomascus nasutus hainanus, el goril·la del riu Cross i l'orangutan de Sumatra.[126]Procolobus badius waldronae fou recentment declarat extingit quan no es trobà cap rastre de la subespècie entre el 1993 i el 1999.[127] Des d'aleshores, alguns caçadors n'han trobat i mort individus i el futur de la subespècie sembla difícil.[128]
Notes
a Els humans habiten a tots els continents si s'inclouen les estacions científiques i meteorològiques a l'Antàrtida.
b Article 6: Tothom i en tot lloc té dret al reconeixement de la pròpia personalitat jurídica.[129]
Referències
↑ Goodman, M., Tagle, D. A., Fitch, D. H., Bailey, W., Czelusniak, J., Koop, B. F., Benson, P., Slightom, J. L. «Primate evolution at the DNA level and a classification of hominoids». Journal of Molecular Evolution, 30, 1990, pàg. 260–266. DOI: 10.1007/BF02099995.
↑ 2,02,12,22,32,42,52,6 «Primate». A: Encyclopædia Britannica Online. Encyclopædia Britannica, Inc., 2008 [Consulta: 21 juliol 2008].
↑ Janečka, J. E.; Miller, W., Pringle, T. H., Wiens, F., Zitzmann, A., Helgen, K. M., Springer, M. S. & Murphy, W. J. «Molecular and Genomic Data Identify the Closest Living Relative of Primates». Science, 318, 5851, 2007, pàg. 792–794. DOI: 10.1126/science.1147555 [Consulta: 17 agost 2008].
↑ Kavanagh, M.. A Complete Guide to Monkeys, Apes and Other Primates. Nova York: Viking Press, 1983, p. 18. ISBN 0670435430.
↑ McKenna, M. C. i Bell, S. K.. Classification of Mammals Above the Species Level. Nova York: Columbia University Press, 1997, p. 329. ISBN 023111012X.
↑ «Nova - Meet Your Ancestors» (en anglès). PBS. [Consulta: 24 octubre 2008].
↑ «Plesiadapis» (PDF). North Dakota Geological Survey. [Consulta: 24 octubre 2008].
↑ Lee, M. «Molecular Clock Calibrations and Metazoan Divergence Dates». Journal of Molecular Evolution, 49, 3, September 1999, pàg. 385–391. DOI: 10.1007/PL00006562.
↑ «Scientists Push Back Primate Origins From 65 Million To 85 Million Years Ago». Science Daily. [Consulta: 24 octubre 2008].
↑ Tavaré, S., Marshall, C. R., Will, O., Soligo, C. & Martin R.D. «Using the fossil record to estimate the age of the last common ancestor of extant primates». Nature, 416, 6882, 18-04-2002, pàg. 726–729. DOI: 10.1038/416726a.
↑ Klonisch, T., Froehlich, C., Tetens, F., Fischer, B. & Hombach-Klonisch, S. «Molecular Remodeling of Members of the Relaxin Family During Primate Evolution». Molecular Biology and Evolution, 18, 2001, pàg. 393–403 [Consulta: 22 agost 2008].
↑ 12,012,112,212,312,4 Horvath, J. et al. «Development and Application of a Phylogenomic Toolkit: Resolving the Evolutionary History of Madagascar's Lemurs» (PDF). Genome Research, 18, 2008, pàg. 490. DOI: 10.1101/gr.7265208 [Consulta: 22 agost 2008].
↑ 13,013,113,213,313,413,5 Error de citació: Etiqueta<ref>no vàlida;
no s'ha proporcionat text per les refs amb l'etiquetaMSW3
↑ 14,014,114,214,314,4 Sellers, Bill. «Primate Evolution» (PDF) p. 13-17. University of Edinburgh, 20-10-2000. Arxivat de l'original el 29 octubre 2008. [Consulta: 23 octubre 2008].
↑ 15,015,115,215,3 Hartwig, W.. «Primate Evolution». A: Campbell, C., Fuentes, A., MacKinnon, K., Panger, M. & Bearder, S.. Primates in Perspective. Oxford University Press, 2007, p. 13–17. ISBN 978-0-19-517133-4.
↑ Ciochon, R. & Fleagle, J.. Primate Evolution and Human Origins. Menlo Park (Califòrnia): Benjamin/Cummings, 1987, p. 72. ISBN 9780202011752.
↑ 17,017,1 Garbutt, N. Mammals of Madagascar, A Complete Guide. A&C Black Publishers, 2007, p. 85–86. ISBN 978-0-300-12550-4.
↑ 18,018,1 Mittermeier, R.A. [et al]. Lemurs of Madagascar. 2a ed.. Conservation International, 2006, p. 23–26. ISBN 1-881173-88-7.
↑ Shekelle, M. (2005). "Evolutionary Biology of Tarsiers". [Consulta: 22 agost 2008]
↑ Schmidt, T. et al. «Rapid electrostatic evolution at the binding site for cytochrome c on cytochrome c oxidase in anthropoid primates». Proceedings of the National Academy of Sciences of the United States of America, 102, 18, 2005, pàg. 6379–6384. DOI: 10.1073/pnas.0409714102 [Consulta: 22 agost 2008].
↑ Marivaux, L. et al. «Anthropoid primates from the Oligocene of Pakistan (Bugti Hills): Data on early anthropoid evolution and biogeography». Proceedings of the National Academy of Sciences of the United States of America, 102, 24, 14-06-2005, pàg. 8436–8441. DOI: 10.1073/pnas.0503469102 [Consulta: 22 agost 2008].
↑ Schrago, C. G. & Russo, C. A. M. «Timing the Origin of New World Monkeys» (PDF Reprint). Molecular Biology and Evolution, 20, 10, 2003, pàg. 1620–1625. DOI: 10.1093/molbev/msg172.
↑ Houle, A. «The origin of platyrrhines: An evaluation of the Antarctic scenario and the floating island model». American Journal of Physical Anthropology, 109, 4, 1999, pàg. 541–559. DOI: 10.1002/(SICI)1096-8644(199908)109:4<541::AID-AJPA9>3.0.CO;2-N.
↑ Andrews, P. & Kelley, J. «Middle Miocene Dispersals of Apes». Folia Primatologica, 78, 2007, pàg. 328–343. DOI: 10.1159/000105148.
↑ 25,0025,0125,0225,0325,0425,0525,0625,0725,0825,0925,1025,1125,1225,1325,1425,1525,1625,1725,1825,1925,20 Strier, K.. Primate Behavioral Ecology. 3a ed.. Allyn & Bacon, 2007, p. 7, 64, 71, 77, 182–185, 273–280, 284, 287–298. ISBN 0-205-44432-6.
↑ Pough, F. W., Janis, C. M. & Heiser, J. B.. «Primate Evolution and the Emergence of Humans». A: Vertebrate Life. 7a ed.. Pearson, 2005, p. 650. ISBN 0-13-127836-3.
↑ Groves, C. P.. Primate Taxonomy. Smithsonian Institution Press, 2001. ISBN 1-56098-872-X.
↑ von Linné, C.. Sistema naturae per regna tria Naturae, secundum classes, ordines, genera, species, cum characteribus differentiis, synonimis locis. Tomus I. Impensis direct. Laurentii Salvii, Holmia, 1758, p. 20–32.
↑ von Linné, C.. Sistema naturae sive regna tria Naturae systematice proposita per classes, ordines, genera, & species. apud Theodorum Haak, Lugduni Batavorum, 1735, p. s.p..
↑ Blainville, H.. «Nouvelle classification des Mammifères». A: Annales Françaises et Etrangères d'Anatomie et de Physiologie Appliquées à la Médicine et à l'Histoire Naturelle, 3, 1839, p. 268–269.
↑ Thorington, R. W., Jr. & Anderson, S.. «Primates». A: Anderson, S. & Jones, J. K., Jr.. Orders and Families of Recent Mammals of the World. Nova York: John Wiley and Sons, 1984, p. 187–217.
↑ McKenna, M. C. & Bell, S. K.. Classification of Mammals: Above the species level. Nova York: Columbia University Press, 1997, p. 631. ISBN 0-231-11013-8.
↑ Strier, K.. Primate Behavioral Ecology. 3a ed., 2007, p. 50–53. ISBN 0-295-44432-6.
↑ Tenaza, R. «Songs of hybrid gibbons (Hylobates lar × H. muelleri)». American Journal of Primatology, 8, 3, 1984, pàg. 249–253. DOI: 10.1002/ajp.1350080307.
↑ 35,035,1 Bernsteil, I. S. «Naturally occurring primate hybrid». Science, 154, 3756, 1966, pàg. 1559–1560. DOI: 10.1126/science.154.3756.1559. PMID: 4958933.
↑ Sugawara, K. «Sociological study of a wild group of hybrid baboons between Papio anubis and P. hamadryas in the Awash Valley, Ethiopia». Primates, 20, 1, 1979, pàg. 21–56. DOI: 10.1007/BF02373827.
↑ Jolly, C. J. et al. «Intergeneric Hybrid Baboons». International Journal of Primatology, 18, 4, 1997, pàg. 597–627. DOI: 10.1023/A:1026367307470.
↑ 38,0038,0138,0238,0338,0438,0538,0638,0738,0838,0938,1038,1138,1238,1338,1438,1538,16 Pough, F. W., Janis, C. M. & Heiser, J. B.. «Characteristics of Primates». A: Vertebrate Life. 7a ed.. Pearson, 2005, p. 630. ISBN 0-13-127836-3.
↑ 39,039,1 Soligo, C., Müller, A.E. «Nails and claws in primate evolution». Journal of Human Evolution, 36, 1999, pàg. 97–114. DOI: 10.1006/jhev.1998.0263.
↑ 40,0040,0140,0240,0340,0440,0540,0640,0740,0840,0940,1040,1140,12 Macdonald, David. «Primates». A: The Encyclopedia of Mammals. The Brown Reference Group plc, 2006, p. 290–307. ISBN 0-681-45659-0.
↑ 41,041,141,2 White, T. & Kazlev, A. «Archonta: Primates». Palaeos, 08-01-2006. [Consulta: 3 juny 2008].
↑ 42,042,142,2 Pough, F. W., Janis, C. M. & Heiser, J. B.. «Primate Societies». A: Vertebrate Life. 7a ed.. Pearson, 2005, p. 621–623. ISBN 0-13-127836-3.
↑ Campbell, B. G. & Loy, J. D.. Humankind Emerging (8a edició). Allyn & Bacon, 2000, p. 85. ISBN 0673523640.
↑ 44,044,1 Aiello, L. & Dean, C.. An Introduction to Human Evolutionary Anatomy. Academic Press, 1990, p. 193. ISBN 0120455900.
↑ 45,045,145,245,345,445,545,6 Myers, P. «"Primates" (On-line)». Animal Diversity Web, 1999. [Consulta: 3 juny 2008].
↑ Ash, M. M., Nelson, S. J. & Wheeler, R. C.. Wheeler's Dental Anatomy, Physiology, and Occlusion. W. B. Saunders, 2003, p. 12. ISBN 9780721693828.
↑ Arrese, C. A., et al. «Cone topography and spectral sensitivity in two potentially trichromatic marsupials, the quokka (Setonix brachyurus) and quenda (Isoodon obesulus)». Proceedings of Biological Science, 272, 1565, 2005, pàg. 791–796. DOI: 10.1098/rspb.2004.3009.
↑ Bowmaker, J. K. & Astell, S. «Photosensitive and photostable pigments in the retinae of Old World monkeys» (pdf). Journal of Experimental Biology, 156, 1991, pàg. 1–19. ISSN: 0022-0949 [Consulta: 16 juny 2008].
↑ 49,049,1 Surridge, A. K.; Osorio, D. «Evolution and selection of trichromatic vision in primates» (en anglès). Trends in Ecology and Evolution, 18, 2003, pàg. 198–205. DOI: 10.1016/S0169-5347(03)00012-0.
↑ Lucas, P. W. & Dominy, N. J. «Evolution and function of routine trichromatic vision in primates». Evolution, 57, 2003, pàg. 2636–2643. DOI: 10.1554/03-168.
↑ 51,051,151,251,351,4 Sussman, R. W.. Primate Ecology and Social Structure, Volume 2: New World Monkeys. Revised First Edition. Needham Heights, MA: Pearson Custom Publishing & Prentice Hall, 2003, p. 77–80, 132–133, 141–143. ISBN 0-536-74364-9.
↑ Ralls, K. «Mammals in Which Females are Larger Than Males». The Quarterly Review of Biology, 51, 2, 1976, pàg. 245. DOI: 10.1086/409310.
↑ Lindstedtand & Boyce «Seasonality, Fasting Endurance, and Body Size in Mammals». The American Naturalist, 125, 1985, pàg. 873. DOI: 10.1086/284385.
↑ Frisch, J. E. «Sex-differences in the canines of the gibbon (Hylobates lar)». Primates, 4, 2, 1963, pàg. 1. DOI: 10.1007/BF01659148.
↑ Kay, R. F. «The functional adaptations of primate molar teeth». American Journal of Physical Anthropology, 43, 2, 1975, pàg. 195–215. DOI: 10.1002/ajpa.1330430207.
↑ Crook, J. H.. «Sexual selection, dimorphism, and social organization in the primates». A: Campbell, B. G.. Sexual selection and the descent of man. Aldine Transaction, 1972, p. 246. ISBN 978-0202020051.
↑ 57,057,1 Cheverud, J. M., Dow, M. M. & Leutenegger, W. «The quantitative assessment of phylogenetic constraints in comparative analyses: Sexual dimorphism in body weight among primates». Evolution, 39, 6, 1985, pàg. 1335–1351. DOI: 10.2307/2408790.
↑ Leutenegger, W. & Cheverud, J. M. «Correlates of sexual dimorphism in primates: Ecological and size variables». International Journal of Primatology, 3, 4, 1982, pàg. 387. DOI: 10.1007/BF02693740.
↑ 59,059,1 Plavcan, J. M. «Sexual dimorphism in primate evolution». American Journal of Physical Anthropology, 33, 2001, pàg. 25–53. DOI: 10.1002/ajpa.10011.
↑ O'Higgins, P. & Collard, M. «Sexual dimorphism and facial growth in papionine monkeys». Journal of Zoology, 257, 2, 2002, pàg. 255–272. DOI: 10.1017/S0952836902000857.
↑ Sussman, R. W.. Primate Ecology and Social Structure Volume 1: Lorises, Lemurs and Tarsiers. Needham Heights, MA: Pearson Custom Publishing & Prentice Hall, 1999, p. 78, 89–90, 108, 121–123, 233. ISBN 0-536-02256-9.
↑ Wrangham, R. W.. «Mutualism, kinship and social evolution». A: Current Problems in Sociobiology. Cambridge University Press, 1982, p. 269–289. ISBN 0521242037.
↑ Fiore, A. D. & Campbell, C. J.. «The Atelines». A: Campbell, C. J., Fuentes, A., MacKinnon, K. C., Panger, M. & Bearder, S. K.. Primates in Perspective. Oxford University Press, 2007, p. 175. ISBN 978-0-19-517133-4.
↑ Bartlett, T. Q.. «The Hylobatidae». A: Campbell, C. J., Fuentes, A., MacKinnon, K. C., Panger, M. & Bearder, S. K.. Primates in Perspective. Oxford University Press, 2007, p. 283. ISBN 978-0-19-517133-4.
↑ Knott, C. D. & Kahlenberg, S. M.. «Orangutans in Perspective». A: Campbell, C. J., Fuentes, A., MacKinnon, K. C., Panger, M. & Bearder, S. K.. Primates in Perspective. Oxford University Press, 2007, p. 294. ISBN 978-0-19-517133-4.
↑ Constable, J. L. et al. «Noninvasive paternity assignment in Gombe chimpanzees». Molecular Ecology, 10, 5, 2001, pàg. 1279–1300. DOI: 10.1046/j.1365-294X.2001.01262.x.
↑ 67,067,167,267,367,4 Rowe, N.. The Pictorial Guide to the Living Primates. Pogonias Press, 1996, p. 4, 139, 143, 154, 185, 223. ISBN 0-9648825-0-7.
↑ Shultz, S. & Thomsett, S.. «Interactions between African Crowned Eagles and Their Prey Community». A: McGraw, W., Zuberbuhler, K. & Noe, R.. Monkeys of Tai Forest, An African Primate Community. Cambridge University Press, 2007, p. 181. ISBN 0-521-81633-5.
↑ 69,069,1 Bshary, R.. «Interactions between Red Colobus Monkeys and Chimpanzees». A: McGraw, W., Zuberbuhler, K. & Noe, R.. Monkeys of Tai Forest, An African Primate Community. Cambridge University Press, 2007, p. 155–170. ISBN 0-521-81633-5. Error de citació: Invalid<ref>tag; name "tai2" defined multiple times with different content
↑ 70,070,1 Stanford, C.. Chimpanzee and Red Colobus : the ecology of predator and prey. Harvard University Press, 1998, p. 130–138, 233. ISBN 0-674-00722-0.
↑ 71,071,1 Boinski, S.. «Social Manipulation Within and Between Troops Mediates Primate Group Movement». A: Boinski, S. and Garber, P.. On the Move : how and why animals travel in groups. University of Chicago Press, 2000, p. 447–448. ISBN 0-226-06340-2.
↑ Boesch, C. & Boesch, H. «Tool Use and Tool Making in Wild Chimpanzees». Folia Primatol, 54, 1990, pàg. 86–99. DOI: 10.1159/000156428.
↑ Westergaard, G. C., et al. «Why some capuchin monkeys (Cebus apella) use probing tools (and others do not)». Journal of Comparative Psychology, 112, 2, 1998, pàg. 207–211. DOI: 10.1037/0735-7036.112.2.207.
↑ de Waal, F. B. M. & Davis, J. M. «Capuchin cognitive ecology: cooperation based on projected returns». Neuropsychologia, 41, 2003, pàg. 221–228. DOI: 10.1016/S0028-3932(02)00152-5.
↑ Paar, L. A., Winslow, J. T., Hopkins, W. D. & de Waal, F. B. M. «Recognizing facial cues: Individual discrimination by chimpanzees (Pan troglodytes) and rhesus monkeys (Macaca mulatta)». Journal of Comparative Psychology, 114, 1, 2000, pàg. 47–60. DOI: 10.1037/0735-7036.114.1.47.
↑ Paar, L. A. & de Waal, F. B. M. «Visual kin recognition in chimpanzees». Nature, 399, 1999, pàg. 647. DOI: 10.1038/21345.
↑ Fujita, K., Watanabe, K., Widarto, T. H. & Suryobroto, B. «Discrimination of macaques by macaques: The case of sulawesi species». Primates, 38, 3, 1997, pàg. 233–245. DOI: 10.1007/BF02381612.
↑ Call, J. «Object permanence in orangutans (Pongo pygmaeus), chimpanzees (Pan troglodytes), and children (Homo sapiens)». Journal of Comparative Psychology, 115, 2, 2001, pàg. 159–171. DOI: 10.1037/0735-7036.115.2.159.
↑ Itakura, S. & Tanaka, M. «Use of experimenter-given cues during object-choice tasks by chimpanzees (Pan troglodytes), an orangutan (Pongo pygmaeus), and human infants (Homo sapiens)». Journal of Comparative Psychology, 112, 2, 1998, pàg. 119–126. DOI: 10.1037/0735-7036.112.2.119.
↑ Gouteux, S., Thinus-Blanc, C. & Vauclair, J. «Rhesus monkeys use geometric and nongeometric information during a reorientation task». Journal of Experimental Psychology: General, 130, 3, 2001, pàg. 505–519. DOI: 10.1037/0096-3445.130.3.505.
↑ Tomasello, M. & Call, J.. Primate Cognition. Oxford University Press US, 1997. ISBN 9780195106244.
↑ Liman, E. R. & Innan, H. «Relaxed selective pressure on an essential component of pheromone transduction in primate evolution» (pdf). Proceedings of the National Academy of Sciences of the United States of America, 100, 6, 2003, pàg. 3328–3332. DOI: 10.1073/pnas.0636123100. PMID: 12631698 [Consulta: 23 juliol 2008].
↑ Egnor, R., Miller, C. & Hauser, M.D.. «Nonhuman Primate Communication». A: Encyclopedia of Language and Linguistics (pdf). 2a ed.. Elsevier, 2004. ISBN 0080442994 [Consulta: 23 juliol 2008].
↑ Milliken, G. W.; Ward, J. P.; Erickson, C. J. «Independent digit control in foraging by the aye-aye (Daubentonia madagascariensis)». Folia Primatologica, 56, 4, 1991, pàg. 219–224. DOI: 10.1159/000156551.
↑ Hiller, C. «Theropithecus gelada». Animal Diversity Web, 2000. [Consulta: 8 agost 2008].
↑ Wright, P.; Simmons, E.; Gursky, S.. «Introduction». A: Wright, P., Simmons, E. & Gursky, S.. Tarsiers Past, Present and Future. Rutgers University Press, 2003, p. 1. ISBN 0-8135-3236-1.
↑ Reed, K. & Fleagle, J. «Geographic and climatic control of primate diversity». Proceedings of the National Academy of Sciences of the United States of America, 92, 17, 15-08-1995, pàg. 7874–7876. DOI: 10.1073/pnas.92.17.7874.
↑ Chapman, C. & Russo, S.. «Primate Seed Dispersal». A: Campbell, C. J., Fuentes, A., MacKinnon, K. C., Panger, M. & Bearder, S. K.. Primates in Perspective. Oxford University Press, 2007, p. 510. ISBN 978-0-19-517133-4.
↑ Long, Y. C., Kirkpatrick, R. C., Zhong, T., and Xiao, L. «Report on the distribution, population, and ecology of the Yunnan snub-nosed monkey (Rhinopithecus bieti)». Primates, 35, 1994, pàg. 241–250. DOI: 10.1007/BF02382060.
↑ Schaller, G. B.. The Mountain Gorilla: Ecology and Behavior. Chicago: University Chicago Press, 1963. ISBN 978-0226736358.
↑ Wolfe, L. D. & Fuentes, A.. «Ethnoprimatology». A: Campbell, C. J., Fuentes, A., MacKinnon, K. C., Panger, M. & Bearder, S. K.. Primates in Perspective. Oxford University Press, 2007, p. 692. ISBN 978-0-19-517133-4.
↑ «Sex is monkey business, too». [Consulta: 30 maig 2009].
↑ Renquist, D. M. & Whitney, R. A. «Zoonoses Acquired from Pet Primates». Veterinary Clinics of North America, Small Animal Practice, 17, 1, 1987, pàg. 219–240 [Consulta: 11 agost 2008].
↑ 94,094,1 Cavalieri, P. & Singer, P. «Declaration on Great Apes». Great Ape Project. [Consulta: 16 juny 2008].
↑ Glendinning, L. «Spanish parliament approves 'human rights' for apes». The Guardian, 26-06-2008 [Consulta: 10 novembre 2008].
↑ Singer, P. «Of great apes and men». The Guardian, 18-07-2008 [Consulta: 10 novembre 2008].
↑ Mager, B. «The HSUS Applauds the U.S. Senate for Unanimously Passing the Captive Primate Safety Act». Humane Society of the United States, 06-07-2006. [Consulta: 10 novembre 2008].
↑ 98,098,1 Workman, C. «Primate conservation in Vietnam: toward a holistic environmental narrative». American Anthropologist, 106, 2, 2004, pàg. 346–352. DOI: 10.1525/aa.2004.106.2.346.
↑ «IPPL News: The US Pet Monkey Trade». International Primate Protection League, 2003. [Consulta: 4 agost 2008].
↑ Bushnell, D. «The Beginnings of Research in Space Biology at the Air Force Missile Development Center, 1946-1952». History of Research in Space Biology and Biodynamics. NASA, 1958. [Consulta: 18 agost 2008].
↑ Blumenthal, D. «Monkeys as Helpers To Quadriplegics At Home». The New York Times, 17-06-1987 [Consulta: 8 octubre 2008].
↑ 102,0102,1 «The supply and use of primates in the EU». European Biomedical Research Association, 1996. [Consulta: 18 agost 2008].
↑ Chen, F. C. & Li, W. H. «Genomic divergences between humans and other hominoids and the effective population size of the common ancestor of humans and chimpanzees». American Journal of Human Genetics, 68, 2, 2001, pàg. 444–456. DOI: 10.1086/318206. PMID: 11170892 [Consulta: 8 octubre 2008].
↑ Conlee, K. M., Hoffeld, E. H. & Stephens, M. L. «A Demographic Analysis of Primate Research in the United States» ( PDF). ATLA (Alternatives to Laboratory Animals), 32, Sup 1, 2004, pàg. 315–322 [Consulta: 8 octubre 2008].
PDF). ATLA (Alternatives to Laboratory Animals), 32, Sup 1, 2004, pàg. 315–322 [Consulta: 8 octubre 2008].
↑ presented to Parliament by the Secretary of State for the Home Department by Command of Her Majesty, July 2006.. Statistics of scientific procedures on living animals: Great Britain 2005 (pdf). The Stationery Office, Juliol 2006. ISBN 0101687729 [Consulta: 16 juny 2008].
↑ «Nonhuman Primates: Research Animals». Animal Welfare Information Center. United States Department of Agriculture. [Consulta: 14 juliol 2008].
↑ «Directive 86/609». European Coalition to End Animal Experiments. [Consulta: 8 octubre 2008].
↑ Grubb, P. et al.. «The Sierra Leone monkey drives». A: Mammals of Ghana, Sierra Leone, and the Gambia (en anglès). St. Ives: Trendrine, 1998, p. 214–219. ISBN 0951256246.
↑ 109,0109,1 Chapman, C. A.; Peres, C. A. «Primate conservation in the new millennium: the role of scientists» (en anglès). Evolutionary Anthropology, 10, 2001, pàg. 16–33. DOI: 10.1002/1520-6505(2001)10:1<16::AID-EVAN1010>3.0.CO;2-O.
↑ 110,0110,1 Mittermeier, R. A.; Cheney, D. L.. «Conservation of primates and their habitats». A: Smuts, B. B., Cheney, D. L., Seyfarth, R. M., Wrangham, R. W.; Struhsaker, T. T.. Primate Societies (en anglès). Chicago: University of Chicago Press, 1987, p. 477–490.
↑ 111,0111,1 Southwick, C. H.; Siddiqi, M. F. «Status, conservation and management of primates in India» (pdf) (en anglès). Envis Bulletin: Wildlife and Protected Areas, 1, 1, 2001, pàg. 81–91 [Consulta: 4 agost 2008].
↑ 112,0112,1112,2112,3 Cowlishaw, G.; Dunbar, R.. Primate Conservation Biology (en anglès). Chicago: University of Chicago Press, 2000. ISBN 9780226116372.
↑ Purvis, A., Gittleman, J. L., Cowlishaw, G. & Mace, G. M. «Predicting extinction risk in declining species». Proceedings of the Royal Society of London, 267, 2000, pàg. 1947–1952. DOI: 10.1098/rspb.2000.1234.
↑ 114,0114,1114,2 Fa, J. E., Juste, J., Perez de Val, J. & Castroviejo, J. «Impact of market hunting on mammal species in Equatorial Guinea». Conservation Biology, 9, 5, 1995, pàg. 1107–1115. DOI: 10.1046/j.1523-1739.1995.9051107.x.
↑ Hill, C. M. «Crop-raiding by wild vertebrates: The farmer's perspective in an agricultural community in western Uganda». International Journal of Pest Management, 43, 1, 1997, pàg. 77–84. DOI: 10.1080/096708797229022.
↑ Hill, C. M. «Primate conservation and local communities: Ethical issues and debates». American Anthropologist, 104, 4, 2002, pàg. 1184–1194. DOI: 10.1525/aa.2002.104.4.1184.
↑ Choudhury, A. «Primates in Northeast India: an overview of their distribution and conservation status» (pdf). Envis Bulletin: Wildlife and Protected Areas, 1, 1, 2001, pàg. 92–101 [Consulta: 4 agost 2008].
↑ Kumara, H. N. & Singh, M. «Distribution and abundance of primates in rainforests of the Western Ghats, Karnataka, India and the conservation of Macaca silenus». International Journal of Primatology, 25, 5, 2004, pàg. 1001–1018. DOI: 10.1023/B:IJOP.0000043348.06255.7f.
↑ Nijman, V. «Conservation of the Javan gibbon Hylobates moloch: population estimates, local extinction, and conservation priorities» (pdf). The Raffles Bulletin of Zoology, 52, 1, 2004, pàg. 271–280 [Consulta: 4 agost 2008].
↑ O'Brien, T. G., Kinnaird, M. F., Nurcahyo, A., Iqbal, M. & Rusmanto, M. «Abundance and distribution of sympatric gibbons in a threatened Sumatran rain forest». International Journal of Primatology, 25, 2, 2004, pàg. 267–284. DOI: 10.1023/B:IJOP.0000019152.83883.1c.
↑ Estrada, A., Coates-Estrada, R. & Meritt, D. «Non-flying mammals and landscape changes in the tropical forest region of Los Tuxtlas, Mexico». Ecography, 17, 1994, pàg. 229–241. DOI: 10.1111/j.1600-0587.1994.tb00098.x.
↑ Marsh, L. K.. «The nature of fragmentation.». A: Marsh, L. K.. Primates in Fragments: Ecology and Conservation. Nova York: Kluwer Academic/Plenum Publishers, 2003, p. 1–10.
↑ Turner, I. M. «Species loss in fragments of tropical rain forest: a review of the evidence». Journal of Applied Ecology, 33, 1996, pàg. 200–209. DOI: 10.2307/2404743.
↑ Chiarello, A.G.. «Primates of the Brazilian Atlantic forest: the influence of forest fragmentation on survival». A: Marsh, L. K.. Primates in Fragments: Ecology and Conservation. Nova York: Kluwer Academic/Plenum Publishers, 2003, p. 99–121. ISBN 9780306476969.
↑ Pope, T.R.. «Socioecology, population fragmentation, and patterns of genetic loss in endangered primates». A: Avise, J. & Hamrick, J.. Conservation Genetics: Case Histories from Nature. Norwell: Kluwer Academic Publishers, 1996, p. 119–159. ISBN 9780412055812.
↑ Mittermeier, R. A. et al. «Primates in Peril: The World's 25 Most Endangered Primates, 2006 – 2008» (pdf). Primate Conservation, 22, 2007, pàg. 1–40 [Consulta: 4 agost 2008].
↑ Oates, J. F., Abedi-Lartey, M., McGraw, W. S., Struhsaker, T. T. & Whitesides, G. H. «Extinction of a West African Red Colobus Monkey». The Journal of the Society for Conservation Biology, 14, 5, 2000, pàg. 1526. DOI: 10.1046/j.1523-1739.2000.99230.x.
↑ McGraw, W. S. «Update on the Search for Miss Waldron's Red Colobus Monkey». International Journal of Primatology, 26, 3, 2005, pàg. 605–619. DOI: 10.1007/s10764-005-4368-9.
↑ «The Universal Declaration of Human Rights». Nacions Unides, 1948. [Consulta: 2 desembre 2008].
Enllaços externs
PrimateImages: Natural History Collection (en anglès)
Primate Info Net (en anglès)
Els primats a l'Animal Diversity Web (en anglès)
Institut d'Investigació dels Priamts de la Universitat de Kyoto (en anglès), (japonès)
Atles citoarquitecturals d'alta resolució del cervell dels primats (en anglès)
EUPRIM-Net: European Primate Network (en anglès)
Ordres vivents de mamífers per infraclasses | |
|---|---|
Regne: Animalia · Embrancament: Chordata · Subfílum: Vertebrata · (sense rang): Amniota | |
| Prototeris | Monotremes |
| Metateris | Didelfimorfs · Opòssums rata · Microbioteris · Talps marsupials · Dasiüromorfs · Peramelemorfs · Diprotodonts |
| Euteris | Afrosorícids · Macroscelidis · Tubulidentats · Damans · Proboscidis · Sirenis · Cingulats · Pilosos · Tupaies · Lèmurs voladors · Primats · Rosegadors · Lagomorfs · Eulipotifles · Ratpenats · Pangolins · Carnívors · Perissodàctils · Artiodàctils |
Famílies vivents de primats per subordre | |
|---|---|
Regne Animalia · Embrancament Chordata · Classe Mammalia · Infraclasse Eutheria · Superordre Euarchontoglires | |
| Strepsirrhini | Cheirogaleidae · Lemuridae · Lepilemuridae · Indriidae · Daubentoniidae · Lorisidae · Galagidae |
| Haplorhini | Tarsiidae · Cebidae · Callitrichidae · Aotidae · Pitheciidae · Atelidae · Cercopithecidae · Hylobatidae · Hominidae |


