Cladística

Aquest cladograma mostra la relació entre diversos grups d'insectes. En alguns cladogrames, la longitud de les línies horizontals indica el temps passat des de l'últim ancestre comú.
La cladística (del grec κλάδος, klados = branca) o sistemàtica filogenètica és una branca de la biologia que defineix les relacions filogenètiques entre els organismes basant-se en similituds derivades, és a dir característiques evolutives noves compartides per un grup d'organismes no presents en d'altres. Per exemple, la presència de columna vertebral i ossos és una novetat evolutiva exclusiva dels vertebrats, i això els defineix com a grup.
És la més important de les sistemàtiques filogenètiques, que estudien les relacions evolutives entre els organismes. La cladística és un mètode d'anàlisi rigorós que utilitza les "propietats derivades compartides" (sinapomorfies: vg. a baix) dels organismes que s'estan estudiant. L'anàlisi cladística forma la base de la majoria dels sistemes moderns de classificació biològica, que busquen agrupar als organismes per les seves relacions evolutives. En contrast, la fenètica agrupa els organismes basant-se en la seva similitud global, mentre que els enfocaments més tradicionals tendeixen a basar-se en caràcters clau. Willi Hennig (1913-1976) és considerat com el fundador de la cladística.
Contingut
1 Cladogrames
2 Definicions
3 Mètodes cladístics
4 Classificació cladística
5 Aplicacions
6 Vegeu també
7 Bibliografia
8 Enllaços externs
Cladogrames

Com a resultat final de l'anàlisi cladística s'obtenen diagrames de relació en arbre anomenats "cladogrames", per mostrar les diferents hipòtesis sobre les relacions. Una anàlisi cladística pot basar-se en tanta informació com l'investigador seleccioni. La investigació sistemàtica moderna sol basar-se en una gran varietat d'informació, incloent seqüències d'ADN (dades moleculars), dades bioquímiques i dades morfològiques.
En un cladograma tots els organismes es col·loquen als extrems de les branques, i cada node interior és idealment binari (amb dues branques). Els dos tàxons de cada bifurcació es diuen taxons germans o grups germans. Cada subarbre, independentment del nombre d'elements que contingui, es diu clade. Tots els organismes d'un grup natural estan continguts en un clade que comparteix un avantpassat comú (un que no comparteixin amb cap altre organisme del diagrama). Cada clade es defineix partint d'una sèrie de característiques que apareixen en els seus membres, però no en les altres formes de les quals ha divergit. Aquestes característiques identificadoras del clade es diuen sinapomorfies (caràcters derivats compartits). Per exemple, les ales anteriors endurides (èlitres) són una sinapomorfia dels coleòpters, mentre que la vernació circinada, o desenrotllar els brots nous, és una sinapomorfia de les falgueres.
Definicions
Apomorfia i plesiomorfia: un estat d'un caràcter que sigui present en els dos grups externs (els parents més propers del grup que no són part del propi grup) i en els avantpassats es diu plesiomorfia (que significa "forma propera", també anomenat estat ancestral). Un estat d'un caràcter que només es dóna en els últims descendents es diu apomorfia (que significa "forma separada", també anomenat estat derivat) per a aquest grup. S'utilitzen els adjectius plesiomòrfic i apomòrfic en lloc de "primitiu" i "avançat" per evitar realitzar judicis de valor sobre l'evolució dels estats dels caràcters, ja que ambdós poden ser avantatjosos en diferents circumstàncies.
Sinapomorfia''' i '''simplesiomorfia: Una sinapomorfia és una apomorfia compartida per un grup monofilètic; una simplesiomorfia és una plesiomorfia compartida per un grup monofilètic.- Es defineixen diversos termes més per descriure els cladogrames i les posicions dels elements dins d'ells. Una espècie o clade és basal d'un altre clade si té més caràcters plesiomòrfics que l'altre clade. Normalment un grup basal és molt pobre en espècies comparat amb un grup més derivat. No es requereix que hi hagi un grup basal. Per exemple, si es consideren les aus i els mamífers junts, cap no és basal de l'altre: ambdós tenen molts caràcters derivats.
- Un clade o espècie situat dins d'un altre clade es pot descriure com niat dins de l'altre clade.
Mètodes cladístics
Una anàlisi cladística s'aplica a un cert conjunt d'informacions. Per organitzar aquesta informació es fa una distinció entre caràcters i estat del caràcter. Si considerem el color de les plomes, aquest pot ser blau en una espècie però vermell en una altra. Per tant, les "plomes vermelles" i les "plomes blaves" són dos estats de caràcter del caràcter "color de les plomes".
Antigament l'investigador decidia quins estats de caràcter eren presents abans de l'últim avantpassat comú del grup d'espècies (plesiomorfies) i quins eren presents en l'últim avantpassat comú (sinapomorfies). Normalment, això es fa considerant un o més grups externs (organismes que no es consideren part del grup en qüestió, però que estan relacionats amb el grup). En caracteritzar les divisions cladístiques, només les sinapomorfies són d'utilitat.
Després es dibuixaven i avaluaven diversos cladogrames possibles. Els clades havien de tenir tantes sinapomorfies com fos possible. L'esperança és que un nombre bastant gran de veritables sinapomorfies serà suficient per superar a qualsevol nombre de simplesiomorfies involuntàries (homoplàsies), causades per evolució convergent (és a dir, caràcters que s'assemblen entre ells per les condicions ambientals o la funcionalitat, no per haver estat heretats d'un avantpassat comú). Un exemple conegut d'homoplàsia per evolució convergent són les ales. Encara que les ales de les aus i els insectes poden assemblar-se superficialment i desenvolupen la mateixa funció, ambdues van evolucionar independentment. Si es puntués com "POSITIU" accidentalment a una au i un insecte per al caràcter "presència d'ales", s'introduiria una homoplasia en el conjunt de dades, la qual cosa podria produir resultats erronis.
Quan sorgeixen possibilitats equivalents, se sol elegir una d'elles basant-se en el principi de parsimònia: la disposició més compacta sol ser la millor hipòtesi sobre les relacions (una variació de la navalla d'Occam). Un altre enfocament, particularment útil en l'evolució molecular, és el de màxima versemblança, que selecciona el cladograma òptim que tingui la major versemblança, basant-se en un model de canvis de probabilitat específica.
Sens dubte, ja no es fa així: els prejudicis de l'investigador són alguna cosa a evitar. Avui dia gran part de l'anàlisi es realitza per programari: a part del programari per calcular els mateixos arbres, hi ha programari estadístic sofisticat per proporcionar una base més objectiva.
La cladística ha tardat un temps a establir-se, i encara hi ha gran debat sobre com aplicar les idees de Hennig al món real. Existeix inquietud sobre el fet que la utilització de conjunts de dades molt diferents (per exemple, característiques estructurals contra característiques genètiques) pot produir arbres molt diferents. Tanmateix, en gran part, la cladística ha demostrat la seva utilitat per resoldre filogènies i s'ha guanyat un ampli suport.
En haver-se simplificat la seqüenciació de l'ADN, les filogènies es construeixen cada vegada més amb l'ajuda de les dades moleculars. La sistemàtica computacional permet l'ús d'aquests grans conjunts de dades per construir filogènies objectives. Així es poden filtrar algunes sinapomorfies veritables de l'evolució paral·lela amb major precisió. Un mètode poderós per reconstruir filogènies és l'ús de marcadors retrotransposó genòmics, que són virtualment lliures d'ambigüitat d'acord amb el coneixement actual (encara que això és una suposició basada en estadístiques i pot no ser cert en algun cas específic, encara que és improbable). Idealment s'haurien de combinar les filogènies morfològiques, moleculars i possiblement d'altres (comportamentals, etc.): cap dels mètodes no és "superior", però tots tenen diferents fonts d'error intrínseques. Per exemple, la veritable convergència de caràcters és molt més comuna en morfologia que en les seqüències moleculars, però les veritables reversions de caràcters solen ocórrer només en aquestes últimes. La datació basada en informació molecular és normalment més precisa que la datació de fòssils, però més carregada d'errors (veure rellotge molecular). Combinant i comparant es poden eliminar molts errors.
Classificació cladística
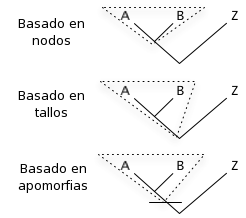
Basat en nodes: l'avantpassat comú més recent d'A i B i tots els seus descendents.
Basat en tiges: tots els descendents de l'avantpassat més vell comú d'A i B que no és avantpassat de Z.
Basat en apomorfies: l'avantpassat comú més recent d'A i B que té una certa apomorfia (caràcter derivat), i tots els seus descendents.
Una tendència recent en la biologia des dels anys 60, denominada o taxonomia cladística requereix que els tàxons siguin clades. En altres paraules, els cladistes argumenten que el sistema de classificació s'ha de reformar per eliminar tot el que no siguin clades. En contrast, altres taxonomistes insisteixen que els grups reflecteixin les filogènies i facin ús freqüent de les tècniques cladístiques, però que es permetin també tàxons parafilètics.
Un grup monofilètic és un clade que comprèn a una forma ancestral i tots els seus descendents, formant així un (i només un) grup evolutiu. Un grup parafilètic és similar, però exclou alguns dels descendents que han sofert canvis significatius. Per exemple, la classe tradicional Reptilia exclou les aus, encara que aquestes van evolucionar a partir d'un rèptil ancestral. Igualment, els invertebrats tradicionals són parafilètics perquè s'exclouen els vertebrats, encara que els últims van evolucionar a partir d'un invertebrat.
Un grup amb membres de línies evolutives separades es diu polifilètic. Per exemple, un grup reconegut antigament, els paquiderms, va resultar ser polifilètic perquè els elefants i els rinoceronts van sorgir separadament d'animals no paquiderms. Tant la taxonomia cladística com l'evolutiva consideren que els grups polifilètics són errors en la classificació que ocorren perquè es va malinterpretar la convergència o alguna altra homoplàsia com una homologia.
Seguint Hennig, els cladistes argumenten que la parafília és igual d'errònia que la polifília. La idea és que els grups monofilètics poden definir-se objectivament, en termes d'avantpassats comuns o de la presència de sinapomorfies. En canvi, els grups parafilètics i polifilètics es defineixen partint de caràcters clau, i la decisió de quins caràcters són d'importància taxonòmica és inherentment subjectiva. Molts argumenten que condueixen a un pensament "gradístic", pel qual els graus "baixos" avancen cap a graus "avançats", i que al seu torn poden conduir a una teleologia. En els estudis evolutius se sol evitar la teleologia perquè implica un pla que no pot demostrar-se empíricament.
Anant més enllà, alguns cladistes argumenten que els rangs dels grups d'espècies són massa subjectius perquè representin cap informació útil, i per tant argumenten que s'haurien d'abandonar. Per aquesta raó, s'han allunyat de la taxonomia linneana cap a una simple jerarquia de clades.
Altres de cladistes argumenten que tots els tàxons són inherentment subjectius, encara que reflecteixin relacions evolutives, ja que els éssers vius formen un arbre essencialment continu. Tota línia divisòria és artificial i crea una secció monofilètica per dalt i una secció parafilètica per sota. Els tàxons parafilètics són necessaris per classificar seccions antigues de l'arbre. També argumenten que els tàxons parafiléticos proporcionen informació sobre canvis significatius en la morfologia dels organismes, en l'ecologia o en la història de la vida; resumint, que tant els tàxons com els clades són nocions valuoses però diferents, amb propòsits diferents. Molts usen el terme monofília en el seu sentit antic, en el qual inclou la parafilia, i usen el terme alternatiu holofília per descriure els clades (monofília en el sentit d'Hennig).
Actualment està en desenvolupament un codi formal per a la nomenclatura filogenètica, el PhyloCode, per al seu ús en la taxonomia cladística. Es pretén que l'utilitzin tant els que volen abandonar la taxonomia linneana com els que volen usar tàxons i clades alhora.
Aplicacions
La cladística no assumeix cap teoria de l'evolució particular, només el coneixement original de la descendència amb modificació. Per això, els mètodes cladístics es poden aplicar, i així s'ha fet recentment, a sistemes no biològics, com determinar les famílies de llenguatges en la lingüística històrica i filiar manuscrits a la crítica textual.
Vegeu també
- Classificació científica
Bibliografia
- de Queiroz, K. & Gauthier, J.A., 1992. Phylogenetic taxonomy. Annual Review of Ecology and Systematics, 23: 449–480.
- de Luna, E., Guerrero, J. A. & Chew-Taracena, T., 2005. Sistemática Biológica: avances y direcciones en la teoría y los métodos de la reconstrucción filogenética. Hidrobiológica, 15(3): 351-370.
- Goloboff, P. A., 1998. Principios básicos de cladística. Sociedad Argentina de Botánica, Buenos Aires.
- Hamdi, H., Nishio, H., Zielinski, R. & Dugaiczyk, A., 1999. Origin and phylogenetic distribution of Alu DNA repeats: irreversible events in the evolution of primates. Journal of Molecular Biology, 289: 861–871.
- Hennig, W., 1960. Elementos de una sistemática filogenética. EUDEBA, Buenos Aires.
- Kitching, I. J., Forey, P. L., Humphries, C. J. & Williams, D. M., 1998. Cladistics, 2nd edn. Oxford University Press, Oxford.
- Patterson, C., 1982. Morphological characters and homology. In: Joysey, K. A. & Friday, A. E. (eds.): Problems in Phylogenetic Reconstruction. Academic Press, London.
- Shedlock, A. M., Okada, N., 2000. SINE insertions: Powerful tools for molecular systematics. Bioessays, 22: 148–160.
- Swofford, D. L., Olsen, G. J., Waddell, P. J. & Hillis, D. M., 1996. Phylogenetic inference. In: Hillis, D. M., Moritz, C. & Mable, B. K. (eds.): Molecular Systematics. Sunderland, MA: Sinauer Associates.
- Wiley, E. O., 1981. Phylogenetics: The Theory and Practice of Phylogenetic Systematics. Wiley Interscience. New York.
Enllaços externs
| A Wikimedia Commons hi ha contingut multimèdia relatiu a: Cladística |
- La Societat Willi Hennig
- Tree of Life Web Project
- Filogenetica.org: Plataforma de divulgació de la comunidad filogenètica
Filogènia | ||
|---|---|---|
| Camps principals | Filogenètica computacional · Filogènia molecular · Cladística |  |
| Conceptes bàsics | Sinapomorfia · Arbre filogenètic · Xarxa filogenètica · Atracció entre branques llargues · Clade · Llinatge fantasma · Models d'evolució de l'ADN | |
| Mètodes d'inferència | Màxima parsimònia · Màxima versemblança · Neighbor-joining · UPGMA · Inferència Bayesiana · Mínims quadrats ordinaris | |
| Temes actuals | PhyloCode · DNA barcoding | |
| -morfia | Simplesiomorfia · Apomorfia · Plesiomorfia · Sinapomorfia · Autoapomorfia | |
| -filetisme | Monofiletisme / Holofiletisme · Parafiletisme · Polifiletisme | |
Viccionari


